quimico
2011-06-16 07:39



I seguenti utenti ringraziano quimico per questo messaggio: ale93, **R@dIo@TtIvO**
quimico
2011-06-16 07:39
La definizione di alcaloidi fu coniata da Meissner nel 1819 e sta a significare simile agli alcali.
Ma cosa si intende per alcaloide?
(1) Sostanza azotata
(2) Azoto in basso stato di ossidazione
(3) Caratteristiche basiche
(4) Attività farmacologica
Il loro ruolo è molteplice:
A. Protezione vs. predatori
B. Regolatori crescita
C. Mantenimento bilancio ionico
D. Riserva di azoto
E. Eliminazione cataboliti azotati
Esistono eccezioni a quanto appena detto per quanto concerne le caratteristiche che un alcaloide deve possedere: ad es. indicina N-ossido e colchicina.
Di norma queste sostanze sono estratte da fonti naturali (piante, organismi marini, funghi, insetti, animali, batteri).
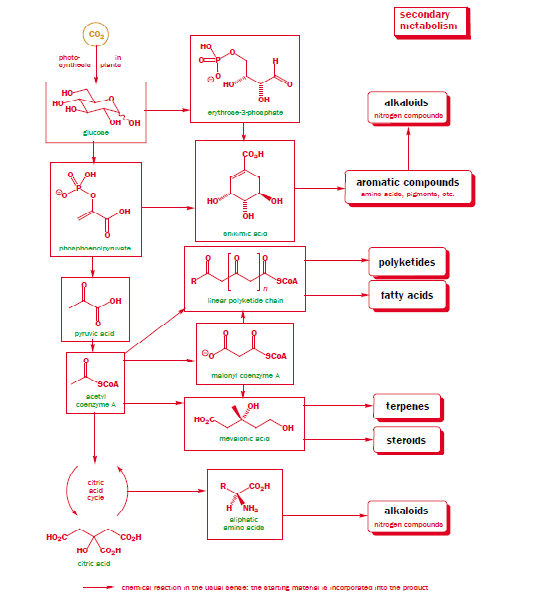
La classificazione è biogenetica (da AA aromatici via acido shikimico e da AA alifatici via ciclo di Krebs):
a) da ornitina (L-Orn)
b) da lisina (L-Lys)
c) da acido nicotinico (da L-Trp)
d) da tirosina (L-Tyr)
e) da triptofano (L-Trp) → alcaloidi indolici
f) da acido antranilico
g) da istidina (L-His)
h) alcaloidi derivanti da amminazione
i) alcaloidi purinici
I seguenti utenti ringraziano quimico per questo messaggio: ale93, **R@dIo@TtIvO**
quimico
2011-06-16 10:03
Tra gli alcaloidi derivanti dall'AA ornitina (di cui sotto riporto la struttura) vorrei citare gli alcaloidi tropanici.
Gli alcaloidi tropanici sono estratti da diverse piante: Atropa belladonna, Datura stramonium, Hyoscyamus niger (Giusquiamo), Mandragora officinarum.
A titolo informativo, come avevo già accennato la sintesi forse più nota del tropinone da cui si ottiene in alcuni passaggi l'atropina è quella biomimetica di Robinson. Immaginò che il tropinone fosse idealmente scomponibile in 3 unità base o sintoni: aldeide succinica, metilamina e acetone; partendo da esse grazie a due condensazioni di Mannich si ottiene infatti con rese scarse il tropinone. La reazione venne migliorata partendo da aldeide succinica, metilamina e, al posto dell'acetone, calcio acetondicarbossilato o etilacetondicarbossilato; one-pot per trattamento finale con HCl si ottiene il prodotto desiderato, ma con una resa del 40%.
In acido acetondicarbossilico, ogni gruppo metilenico è vicino a due gruppi carbonilici, così è per la maggior parte in forma di enolo. Schöpf et al. nel 1935 ottennero una resa del 70-85% conducendo la reazione a pH = 7.
Elming et al. nel 1958 sintetizzarono il tropinone usando metilamina idrocloruro, acido acetondicarbossilico, e generando l'aldeide succinica in situ trattando con acido il 2,5-dimetossitetraidrofurano. La resa fu dell'81%, ma in questo caso non furono necessarie condizioni fisiologiche.
Willstätter sintetizzò il tropinone per altra via:

anche se il suo fine era farsi la cocaina.
Comunque ottenuto il tropinone, questo viene ridotto a tropina, che può essere racema o enantiomericamente pura. La prima volta vennero separate come sali di oro, portando alla tropina che è l'isomero con l'OH assiale, e alla Ψ-tropina che ha l'OH equatoriale.
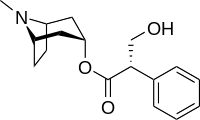
Come detto facendo reagire la tropina con l'acido tropico si ottiene un composto achirale (possiede un piano σ!) noto come iosciamina, la quale a t.a. tautomerizza a (+)-iosciamina e per riscaldamento tautomerizza a (-)-iosciamina.
L'atropina che trova largo uso in ambito oftalmico (sia come midriatico che come cicloplegico) ma anche in altro (ad esempio per la bradicardia o l'avvelenamento da composto organofosfato) è una miscela di iosciamina racema, e in particolare è somministrata sottoforma di solfato monoidrato.
I seguenti utenti ringraziano quimico per questo messaggio: jobba
Max Fritz
2011-06-16 10:37
Ottimo contributo!
Guarda guarda che bello quell'anello con 5 atomi di zolfo consecutivi della varacina... veramente certi esserini a volte umiliano i nostri sforzi nel sintetizzare molecole ben più semplici!!
Ma restando in tema di molecoline interessanti, vorrei chiedere una precisazione su una definizione. La patulina, o anche la cantaridina, che sono due molecole tossiche contenenti solo C, H e O (di origini molto diverse ovviamente), non possono essere definite alcaloidi appunto perchè non contengono N. Ma possono essere definite tossine? O quest'ultima definizione è esclusiva di molecole proteiche?
quimico
2011-06-16 11:06
esistono le eccezioni infatti... alcaloidi è un termine generico.
sinceramente non saprei risponderti. ci vorrebbe DMZ.
più avanti aggiungerà altre chicce 
NaClO3
2011-06-16 11:09
Max, la tossina ha prerogativa di essere di origine biologica, almeno così la definisce il mio libro di Chimica degli alimenti, non sempre è proteica.
quimico
2011-06-18 10:58
Tra gli alcaloidi da tirosina vorrei citare per prime le fenetilamine. Sebbene molte molecole di questa famiglia abbiano assunto una connotazione illegale, io penso che a scopo puramente informativo siano interessanti e da conoscere. Tanto non approfondirò niente e citerò quello che si può trovare su libri. Ho ampia conoscenza di queste molecole, dal punto di vista sintetico ovvio... e ho ben due libri di sole sintesi/ricette ma non sono materia di questo post.
Detto questo, come ho accennato sopra derivano dalla L-Tyr: tra esse cito dopamina (che non è altro che la tirosina decarbossilata, a cui è aggiunto un gruppo OH in posizione 3), la noradrenaliana (che presenta un OH sul carbonio della catena esociclica, a sterochimica R) e l'adrenalina (che è l'analogo NHMe della noradrenalina... il prefisso nor sta ad indicare l'assenza di un gruppo metile).
Al di fuori di questo gruppo di molecole, cito unicamente e a titolo conoscitivo la mezcalina (allucinogeno, estratto dal Peyote o Lophophora sp., 300-500 mg).

La dopamina e la noradrenalina sono neurotrasmettitori nei mammiferi; l'adrenalina nell'uomo. Sono anche note come catecolamine, per l'analogia strutturale col catecolo.
L'adrenalina viene rilasciata dal surrene in situazione di emergenza; stimolazione del catabolismo del glicogeno nel muscolo; aumento della respirazione con produzione di energia mitocondriale (ATP).
La L-DopA (struttura è riportata sopra) è uno, se non il più noto, dei farmaci creati per la cura del Morbo di Parkinson (deficienza di dopamina); negli ultimi anni si è vista però la comparsa di quella che è stata definita "Sindrome da L-DopA" ovvero, dopo 4-5 anni dall'assunzione di tale farmaco, nel 50-80% dei casi si ha la comparsa di anoressia, pressione arteriosa bassa, turbe psicotiche...).
La prima sintesi di tale molecola partiva direttamente da L-Tyr, la quale per ossidrilazione enzimatica veniva converita in L-DopA (è stata abbandonata per le rese troppo basse, inaccettabili). Un'altra sintesi che partiva sempre da L-Tyr prevedeva l'uso della reazione di Fenton (H2O2/FeSO4/hν) venne anch'essa abbandonata per le rese troppo basse e la regiochimica scarsa.
Si decise così di partire da molecole diverse che possedevano già parte del corredo chimico necessario per l'ottenimento del prodotto; si scelsero così vanillina e piperonalio.
Vediamo ora i due casi:
- vanillina

- piperonalio

quimico
2011-06-19 10:20
Continuando la carrellata degli alcaloidi derivati dalla L-Tyr non posso che citare le tetraidroisochinoline, e in particolare la ben nota papaverina, oppiaceo usato come vasodilatatore, spasmolitico; nel papavero da oppio (P. somniferum) si può trovare lo 0.5-2.5% di tale alcaloide. Nell'oppio il 25% ca. è costituito da alcalolidi (ca. 40 alcaloidi identificati), e tra essi si annoverano: morfina (4-21%), codeina (0.8-2.5%), tebaina (0.5-2%), la già citata papaverina, narcotina (4-8%) e narceina (0.1-2%), per citare le più note e importanti. Normalmente si trovano sottoforma di meconati o esteri dell'acido meconico.
All'interno dell'oppio ci sono quindi 2 classi di alcalodi:
- acidi (morfina, narceina);
- basici (codeina, tebaina, narcotina, papaverina).
Sorvolo sul procedimento di estrazione per ovvi motivi.
Nelle regioni note come Triangolo d'oro e Mezzaluna d'oro si era cercato di porre fine al genere P. somniferum inserendo il genere P. bactreatum ma è stato un insuccesso.
Tornando a noi, come dicevo, gli alcaloidi sopra citati hanno tutti come biogenesi l'aminoacido L-Tyr e vengono biosintetizzati via (S)-reticulina; questa viene ossidata a molecola achirale per poi esser ridotta sempre enzimaticamente a isomero (R)-, la forma necessaria a questa biosintesi.
Dopo un'ossidazione enzimatica cui segue accoppiamento omolitico si ottiene la salutaridina, che viene subito ridotta ad alcole bis-allilico, acilata tramite CoA2S e spontaneamente chiude a tebaina.

Dalla tebaina si ottengono tutta una serie di molecole importanti di cui parleremo dopo.
Ora vorrei solo fare una considerazione su quelli che vengono chiamati morfinani. Derivano dalla morfina e si ottengono per demolizione successiva della molecola, che idealmente è come se perdesse gli anelli che formano la molecola. La dizione morfinani è intesa sia in generale sia in particolare.
In particolare i morfinani più noti sono il destrometorfano (antitussivo) e il levallorfano (Lorfan, potente morfino-antagonista).
Abbiamo poi i benzomorfani, tra cui ricordo solo la pentazocina (usato in ostetricia, non deprime il respiro del neonato, cosa che invece fa la morfina. Poi le fenilpiperidine tra cui cito solamente la petidina (analgesico più blando della morfina, con minori effetti collaterali) e il fentanil (analgesico 100x più potente della morfina) e infine abbiamo le fenilpropilamine tra cui ricordo il metadone. Molecola che si può anche sintetizzare in laboratorio, ma anche qui per ovvi motivi non vi dico niente.
Tornando sul tema tebaina, vi mostro le modificazioni strutturali di essa che danno vita a diverse molecole usate soprattutto in ambito farmacologico, con la sola eccezione della eroina (anche se nel 1800 fu usata ampiamente come principio attivo nello sciroppo per la tosse):

Ovviamente dalla tebaina si ricavano molti altri alcaloidi degni di nota, uno su tutti è la morfina.

Concludo la serie citandovi l'ultimo derivato della tebaina, l'etorfina, anestetico troppo potente per esser usato sull'uomo in quanto andrebbe dosato in nM/kg, ha trovato impiego come anestetico per elefanti.

I seguenti utenti ringraziano quimico per questo messaggio: al-ham-bic, Max Fritz, **R@dIo@TtIvO**
Max Fritz
2011-06-19 17:50
Giusto per evitare che sorgano problemi:
quanto scritto da quimico nei post precedenti è di interesse ESCLUSIVAMENTE teorico.
Chiunque venisse qui a chiedere precisazioni su sintesi di alcaloidi (specie se illeciti) verrà severamente punito.
Questo non per criticare il lavoro di quimico, che invece apprezzo molto ovviamente 
I seguenti utenti ringraziano Max Fritz per questo messaggio: jobba
quimico
2011-06-20 07:12
Giusta precisazione Max. Anche io ho sottolineato che sono solo a scopo teorico queste cose. Lungi da me spiegare o dare ricette su queste cose. Sono "solo" interessanti molecole biologicamente attive. E tali restano. Grazie dell'apprezzamento. Prossimamente deciderò se mettere qualcosa sugli alcaloidi indolici monoterpenici.
quimico
2011-06-28 19:40
I Solanum glicoalcaloidi sono metaboliti secondari formati dagli stessi precursori degli steroidi. Colesterolo, colesteranolo, e cicloartenolo sono precursori alternativi per le biosintesi degli agliconi. Gli agliconi hanno lo scheletro steroideo del colestano (C27). L'azoto è adattato ai derivati del colesterolo dagli aminoacidi glicina, arginina (Jadhav and Salunkhe, 1973; Jadhav et al., 1973), o L-arginina (Kaneko et al., 1976).
Gli agliconi sono divisi in cinque differenti categorie a seconda della loro struttura: solanidani hanno anelli indolizidinici fusi, spirosolani hanno una porzione alcaloide ossaazaspirodecanica, 22, 26-epiminocolestani,
α-epiminocicloemichetali, e 3-aminospirostani. La maggior parte dei glicoalcaloidi trovato nelle specie Solanum derivano da solanidani e spirosolani.
I più comuni solanidani sono la solanidina e la sua forma deidrogenata, la demissidina.
Kuhn et al. (1955 a, b) mostrò che la “solanina”, che era stata scoperta nella patata nel 1820, era una miscela di due differenti glicoalcaloidi, α-solanina e α-caconina, entrambe con la solanidina come aglicone ma recanti un differente carboidrato ciascuna. I glicoalcaloidi α-solanina e α-caconina sono generalmente presenti in tandem nelle piante, specialmente in S. tuberosum. L'epimero 22R, 25R della solanidina è stato ritrovato nei tuberi di S. vernei, ma questo ritrovamento non è stato riportato altrove (van Gelder e Scheffer, 1991). La tomatidina è l'aglicone spirosolanico dell'α-tomatina
presente nel pomodoro (Lycopersicon esculentum) così come in S. brevidens, e S. acaule, tra le altre specie. La deidrotomatidina, che è la tomatidina 5,6-deidrogenata, è spesso ritrovata in tandem la struttura della tomatidina, specialmente nelle piante di pomodoro. Le coppie 22, 25-diastereoisomeriche di tomatidina e deidrotomatidina, soladulcidina e solasodina rispettivamente, sono state trovate in molte specie Solanum. Venne quindi suggerito che solanidani e spirosolani sono formati attraverso la stessa via biosintetica fino agli ultimi step, dove la etiolina è trasformata in altri tipi di agliconi (Kaneko et
al., 1976; Petersen et al., 1993). Petersen et al. (1993) suggerirono che gli spirosolani diastereoisomerici sono formati attraverso differenti cammini. Immediatamente dopo la formazione degli agliconi sono glicosilati attraverso gli enzimi glucosiltransferasi (Stapleton et al., 1991).
I saccaridi attaccati alla posizione 3-idrossi degli agliconi consistono di differenti combinazioni di D-glucosio (Glc), D-galattosio (Gal), D-xilosio (Xyl), e L-ramnosio (Rha) nella forma di tetra o trisaccaridi.
Minori quantità di agliconi attaccati a di e monosaccaridi, agliconi liberi, e i loro prodotti dienici, sembrano apparantemente artifatti, e furono rilevati in estratti di patata (Osman and Sinden, 1977; Nikolic and Stankovic, 2003). Il trisaccaride della α-solanina è chiamato solatrioso, e quello dell'α-caconina, un cacotrioso. Un tetrasaccaride, il licotetraoso, è presente nell'α-atomatina ma anche nella demissina, e nella soladulcina B . Inoltre, un altro tetrasaccaride il commertetraoso è legato alla solanidina, producendo la commersonina in S. commersonii, e alla tomatidina, producendo la sisunina in S. acaule x S. ajanhurii. Infatti, questi quattro saccaridi sono i più comuni della vasta varietà dei glicoalcaloidi Solanum.
Se i sei maggiori agliconi sono combinati con i quattro diversi saccaridi, 24 differenti glicoalcaloidi sono ottenuti. Molti di loro sono stati identificati in piante del genere Solanum della sezione Petota. Comunque, alcune combinazioni non sono state riportate, ad es. tomatidina legata a solatriosi, in qualsiasi pianta. In aggiunta a questo, più di 100 specie Solanum sono state analizzate per i glicoalcaloidi α-solanina, α-caconina, α-tomatina, demissina, solamargina, e solasonina (Schreiber, 1968).
I glicoalcaloidi sono composti potenzialmente tossici che hanno un ruolo nel sistema di protezione della pianta. La loro tossicità è basata sulla loro attività anti-colinesterasi sul SNC, sulla rottura delle membrane cellulari attraverso complessazione con i 3β-idrossisteroli nella membrana, e sui cambiamenti causati nel trasporto di ioni attraverso le membrane, col risultato di provocare disordini nel metabolismo generale del corpo (Friedman et al., 1992a; Keukens et al., 1995; Blankemeyer et al., 1992; Blankemeyer et al., 1995).
I sintomi tossici negli umani includono disordini neurologici e gastrointestinali, quali vomito, dolori allo stomaco, tachicardia, e allucinazioni.
La patata coltivata S. tuberosum contiene principalmente α-solanina e α-caconina in rapporto di 0.3 a 0.8 (α-solanina
a α-caconina) (Friedman, 2003). Poichè l'α-caconina è più tossica dell'α-solanina, un rapporto maggiore di α-solanina rispetto all'α-caconina è preferito, e il rapporto dipende dalla coltivazione. Quantità minori di solamarine sono state anche trovata nel fogliame e in tuberi sia vecchi che feriti (Shih and Kuc, 1974; Sinden and Sanford, 1981; Chivanov et al., 2001).
I glicoalcaloidi sono isolati dal materiale della pianta attraverso estrazione con solvente preferibilmente da materiale della pianta asciutto, o in alternativa da materiale fresco. L'asciugatura del materiale della pianta è condotto all'aria o nel forno. Comunque, matariale asciutto congelato può anche essere utilizzato. I materiali freschi sono generalmente raccomandati per essere trasferiti in etanolo per prevenire l'imbrunirsi a causa degli enzimi (Bergers, 1980). In accordo con Dao e Friedman (1996), i risultati delle analisi dei glicoalcaloidi da polveri di foglie asciutte e congelate sono molto più riproducibili di quelli ottenuti da foglie fresche. I principali vantaggi di usare materiale asciutto congelato nell'analisi di glicoalcaloidi sono (1) induzione ridotta, catalizzata da enzimi, di ferite e cambiamenti nella composizione indotti dall'umidità che potrebbero cambiare il contenuto di glicoalcaloidi, e (2) più facile trasporto di campioni per l'analisi in tempi differenti e attraverso diversi ricercatori (Dao and Friedman, 1996). Nessuna degradazione di glicoalcaloidi in campioni asciutti congelati è stata riportata.
I glicoalcaloidi sono solubili in soluzioni acquose acide e in solventi organici polari, inclusi acetonitrile, metanolo, etanolo, e propanolo. In molti casi l'estrazione di glicoalcaloidi da materiali vegetali è condotta in acido acetico diluito (15%), un solvente non tossico e poco costoso.
I solventi organici come metanolo (Fitzpatrick and Osman, 1974) e metanolo miscelato con cloroformio (Wang et al., 1972) sono stati preferiti specialmente per campioni freschi. È stato riportato, comunque, che solventi acquosi sono più efficienti per materiali asciutti che quelli non acquosi (Bushway et al., 1986; Friedman and McDonald, 1995). Combinazioni di differenti solventi sono stati preparate per ottenere estrazione efficace di glicoalcaloidi amfifilici (Friedman and McDonald, 1997, 1999). La possibile idrolisi è evitata usando acidi deboli diluiti o solventi organici a rt. Comunque, metanolo acquoso contenente l'1% di HC è stato anche utilizzato (Kozukue et al., 1999; Stobiecki et al., 2003). Il bisulfito è spesso aggiunto per prevenire l'ossidazione dell'estratto (Hellenäs, 1986; Edwards and Cobb, 1996).
Dopo estrazione, i glicoalcaloidi sono o precipitati con ammoniaca (Fitzpatrick and Osman, 1974; Bushway et al., 1979; Dao and Friedman, 1996; Sotelo and Serrano, 2001; Kozukue and Friedman, 2003), o isolati tramite estrazione in fase solida (SPE) (Carman et al.,
1986; Bushway et al., 1986; Jonker et al., 1992; Friedman et al., 1994; Abell and Sporns, 1996; Edwards and Cobb, 1996; Friedman et al., 1998a; Esposito et al., 2002), o una combinazione di questi metodi. Dopo precipitazione con ammoniaca a pH ~10, il precipitato asciutto di glicoalcaloide è disciolto in metanolo o qualche altro solvente utile al caso, o nuovamente estratto con butanolo (Sotelo and Serrano, 2000). Poiché α-caconina era presente in fase liquida ad alti pH, il metodo di precipitazione può non dare risultati quantitativi (Gregory et al., 1981). Minore recupero con metodo di precipitazione supportato usando metodo SPE per α-tomatina (Friedman et al., 1994).
I seguenti utenti ringraziano quimico per questo messaggio: jobba
jobba
2011-06-28 19:51
Grazie.
quimico
2011-06-29 11:08
Per completezza allego due immagini:
- nella prima sono riportati i tri- e tetrasaccaridi citati sopra;
- nella seconda sono riporati gli alcaloidi.
Unendo le due parti si ottengono i glicoalcaloidi della specie Solanum

 Sperando siano di vostro gradimento e utilità.
Saluti.
Sperando siano di vostro gradimento e utilità.
Saluti.
I seguenti utenti ringraziano quimico per questo messaggio: jobba